

Se divide a su vez en:
Fecundación
Ocurre en el tercio externo de la tuba, y corresponde a la fusión del gameto masculino con el femenino, para formar el cigoto, que experimentará una serie de transformaciones al interior de la tuba.
Segmentación
A las 30 horas el cigoto sufre su primera división mitótica, originando 2 blastómeros. Cuando existen entre 12- 16 blastómeros (después de la 3ª segmentación) se produce un proceso denominado compactación que divide este grupo de células en una zona interior (embrioblasto) y una zona exterior (trofoblasto). El embrioblasto formará al embrión. El trofoblasto formará a los anexos embrionarios.
Al ingresar el conglomerado de células en la cavidad del útero, comienza a entrar líquido a través de la zona pelúcida hacia los espacios intercelulares del embrioblasto, dejando a éste en una posición polar.
En el momento que todo el líquido ingresado se ubica en una cavidad única se le denomina blastocisto.

Implantación
Tras perder la zona pelúcida, en un proceso conocido como Eclosión, el blastocisto de adhiere a la mucosa uterina el 6º día, para estar completamente implantado el día 14.
Pregastrulación
En esta etapa, las células del embrioblasto se ordenan en dos estratos, que se diferencian por su ubicación y forma en: epiblasto e hipoblasto.
Gastrulación (3° semana)
Se inicia con la formación de la línea primitiva (día 15) en el epiblasto, la cual en su extremo anterior o craneal presenta el nódulo o fosita primitiva, que es la entrada hacia un conducto, llamado conducto neuroentérico, que se proyecta hacia anterior, uniendo ambas cavidades (cavidad del saco vitelino y cavidad amniótica).
La gastrulación es el proceso en el que las células del epiblasto, próximas a la línea primitiva, comienzan a proliferar y a penetrar por ella. Algunas células:
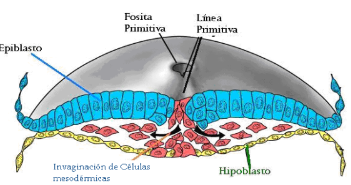
Las células (que no se invaginan) que forman el ectodermo, están organizadas en 3 zonas:
Formación de la notocorda
Existen 2 zonas, en las que no hay una hoja intermedia (mesodermo), que se denominan: membrana bucofaríngea (hacia cefálico) y membrana cloacal (hacia caudal).
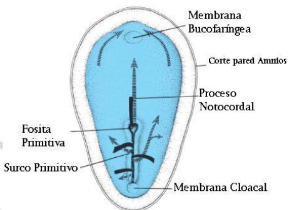

La notocorda definitiva se forma gracias a que las células de la placa notocordal proliferan y se desprenden del endodermo, creando un cordón macizo. El rol de la notocorda es ser inductora de la formación del sistema nervioso (a comienzos de la 3ª semana de desarrollo), a través de moléculas que actúan sobre células del ectodermo, transformándolas en células neuroectodérmicas, comenzando así los mecanismos reguladores de la morfogénesis del tubo neural. La notocorda ha sido capaz de inducir células precursoras del sistema nervioso en otras zonas, lo cual demuestra su alta capacidad inductora. También se han producido malformaciones muy graves al sacarla.
El tejido embrionario que queda alrededor de la notocorda va a formar la vértebra y la notocorda va a formar la parte central del cuerpo de la vértebra. En el adulto existen restos de notocorda a nivel del núcleo pulposo de los discos intervertebrales.
Neurulación
Se le denomina neurulación al proceso en el que se forma el tubo neural y emigran las crestas neurales. Al comenzar la 3ª semana, la notocorda en desarrollo y el mesodermo adyacente estimulan al ectodermo que está encima de ellos. Este complejo proceso de inducción notocordal hace que tejido ectodérmico (neuroectoblasto) se engrose, formándose así la placa neural.
Alrededor del 18º día de desarrollo los bordes laterales de la placa neural se elevan y forman los pliegues neurales; la porción media entre los pliegues neurales forma el surco neural. Hacia el final de la 3ª semana los pliegues neurales se elevan aún más, se acercan y se fusionan irregularmente en la línea media formando el tubo neural. La fusión empieza en la región cervical y sigue hacia cefálico y caudal.

Mientras ocurre la fusión, los bordes libres del ectodermo superficial se separan del tubo neural. Posteriormente, ambos bordes se unen y forman una capa continua en la superficie que dará origen al epitelio epidérmico (epidermoblasto). El punto específico en el que se inicia el contacto y fusión de los pliegues neurales se denomina punto nucal, se ubica entre el 4º y 5º somito. Debido a que la fusión de los pliegues neurales no ocurre simultáneamente a lo largo de ellos, la luz del tubo neural comunica con la cavidad amniótica en sus extremos cefálico y caudal a través de los neuroporos craneal (anterior) y caudal (posterior). El cierre del neuroporo craneal se realiza en ambas direcciones desde el sitio inicial de cierre en la región cervical o desde otro que se origina un tiempo después en el prosencéfalo que avanza en dirección caudal. Este cierre ocurre al 25º día (período 18-20 somitos). El neuroporo caudal se cierra el 27º día (período de 25 somitos). El cierre de ambos neuroporos coincide con el establecimiento de la circulación sanguínea hacia el tubo neural. Un defecto en el cierre de los neuroporos produce una alteración grave en el desarrollo del sistema nervioso (anencefalia y mielosquisis, por ejemplo).
Mientras los pliegues neurales se acercan a la línea media para fusionarse, un grupo de células neuroectodérmicas ubicadas en la cresta de cada pliegue (cresta neural) pierden su afinidad epitelial con las células de la vecindad. Las crestas neurales darán origen a un conjunto heterogéneo de tejidos de gran importancia: ganglios de la raíz posterior, ganglios autónomos, ganglios de los pares craneales V, VII, IX, X, células de schwann, leptomeninges (aracnoides y piamadre , melanocitos, médula suprarrenal, odontoblastos.

En consecuencia, el tubo neural será el que se convertirá por diferenciación en encéfalo y médula espinal, mientras que las crestas neurales formarán la mayor parte del sistema nervioso periférico (SNP) y parte del autónomo (SNA). Una vez formado el tubo neural queda con una cavidad en su interior que en el adulto, permanece en el cerebro a nivel de los ventrículos laterales y tercer ventrículo; en el tronco encefálico en el cuarto ventrículo y en la médula como el canal central de la médula.

1. Inducción
La inducción es el mecanismo por el cual la actividad de un tejido es capaz de determinar o modificar la actividad o destino de otro. La notocorda ejerce un efecto inductor sobre las células ectodérmicas determinando la formación de la placa neural. La zona que queda por encima de la notocorda se denomina encéfalo epicordal. A nivel celular (de las células de la placa neural) existe un aumento de microtúbulos, microfilamentos en la zona apical de las células, permitiendo cambios conformacionales, pasando de células aplanadas a células cilíndricas.
La separación del tubo neural está mediada por la presencia de Moléculas de Adhesión Celular (MAC), del tipo E-Cadherinas, que son las primeras en aparecer, posterior a la inducción se comienzan a expresar las N-cadherinas y N-MAC por lo que estos tejidos después no se vuelven a adherir nunca más, es decir, el tubo neural se separa de las células ectodérmicas definitivamente.
Etapas de Regionalización
Posteriormente, existe una etapa de regionalización del sistema nervioso, mediada por la acción de sustancias químicas (factores neuralizantes y regionalizantes), que actúan sobre el genoma de células ectodérmicas. Una vez cerrado el tubo, se regionaliza diferenciándose:
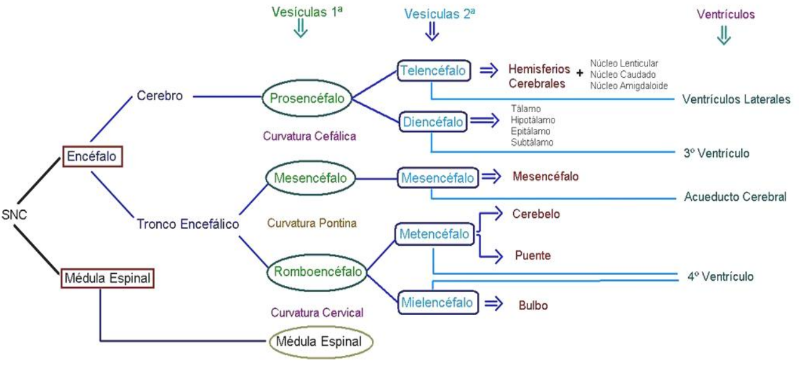
Al término de la 3° semana vamos a encontrar un embrión que presenta en su tubo neural 3 vesículas y 2 flexuras o curvaturas (cefálica y cervical).
La curvatura cefálica es la primera en aparecer, se ubica entre el prosencéfalo y el mesencéfalo.. Se produce cuando placa precordal y la notocorda dejan de sustentar al tubo neural, provocando que la porción que queda afuera de este sustento se caiga, produciéndose así la primera curvatura del tubo neural, la curvatura cefálica. (Recordar que la placa precordal y la notocorda, se extienden por la línea media debajo del ectodermo, desde cefálico a caudal a modo de eje que sirve de soporte y guía al tubo neural).
La curvatura cervical aparece entre el romboencéfalo y la médula.
Estas son las 1ª curvaturas que aparecen, y eso le da un aspecto característico al embrión que ha crecido mucho en la parte rostral o cefálica y que se ha ido acodando debido al crecimiento mayor en la parte dorsal respecto de la ventral. A pesar de todas las acodaduras siempre va a existir un tubo y una cavidad interna.
Al termino de la 4° semana y principios de la 5° aparece una tercera curvatura, la curvatura pontina, pero es inversa a las anteriores (hacia dorsal, lo que señala un mayor crecimiento en la región ventral del embrión).
La curvatura pontina aparece entre mesencéfalo y romboencéfalo.
En la 5° semana aparece se subdividen 2 de las vesículas primarias, para originar un embrión con 5 vesículas secundarias.

Las vesículas secundarias más la médula espinal van a dar origen a todas las partes del sistema nervioso
En el proceso de regionalización se postula la existencia de genes muy específicos denominados genes homeóticos, los cuales serían responsables de regular la regionalización céfalo-caudal del individuo, éstos se ordenan en los cromosomas en la misma secuencia que se expresan en el eje céfalo-caudal del organismo: los genes encargados de la estructura anterior son los genes 3’, en tanto que los genes 5’ se encargan de las estructuras caudales o posteriores.
Se ha postulado que el ácido retinoico, como un morfógeno, actuaría sobre los genes homeóticos modificando su expresión.
2. Proliferación
La proliferación ocurre cuando el tubo neural esta constituido por un epitelio de aspecto pseudoestratificado, cuyas células conectan sus extremos apical y basal a las membranas limitantes externas e internas. Durante esta etapa se observan las siguientes dinámicas celulares:
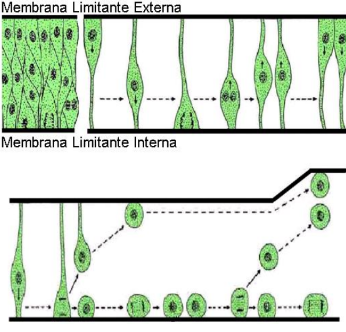
La duplicación del ADN se realiza mientras el núcleo viaja hacia la M.L.E., ocurriendo todo este proceso en la interfase. Termina a nivel de la M.L.E., desde donde el núcleo vuelve nuevamente hacia la M.L.I. y comienza a encogerse lentamente. Su citoplasma se retrae y su núcleo va avanzando hasta que llega a convertirse en una célula redondeada y en ese momento ocurre el proceso de separación de las dos células hijas.
Cuando el eje de separación es vertical quedan dos células que siguen adheridas a la membrana limitante interna. Por lo tanto estas dos células van a seguir siendo células precursoras, que siguen el mismo proceso. De esta forma se asegura la formación de gran cantidad de neuroblastos.
Si el eje de separación es horizontal una célula queda adherida a la M.L.I. y la otra queda libre, ésta se desprende y sale hacia fuera, hacia la Capa del Manto, que es la capa celular que queda inmediatamente alrededor del canal central (que contiene cuerpos neuronales, la sustancia gris).
Por fuera de la Capa del Manto existe otra capa que es la Capa Marginal, que corresponde a las prolongaciones dendríticas y axónicas provenientes de los neuroblastos de la capa de manto. Por lo tanto la capa marginal va a dar origen a la sustancia blanca.
Grandes cantidades de neuronas aparecen desde el tercer trimestre de gestación, hasta el primer año de vida postnatal, debido a la diferenciación del neuroepitelio, el cual produce neuroblastos (células totipotenciales), que pasan por etapas apolar, bipolar, multipolar, hasta llegar a neuronas maduras.
El neuroepitelio, lo primero que produce son los neuroblastos, y cuando dejan de producirlos, comienzan a producir células cuya función es distinta a la de la neurona.

Un tipo celular formado son los glioblastos, que son precursores de tres tipos de células propias del sistema nervioso: astrocitos protoplasmáticos, astocitos fibrosos y oligondedrocitos. Los 2 primeros son muy importantes en la constitución de la barrera hematoencefálica.
Una vez que se han producido los glioblastos, las células que quedan definitivamente allí son los ependimocitos o células ependimarias, que son células que cubren todos los espacios alrededor de las cavidades del sistema nervioso.
Alrededor del 4º mes aparecen las células de microglia, que no tienen origen ectodérmico. Derivan del mesénquima circundante y se caracterizan por ser pequeñas y muy fagocíticas. Llegan a la sustancia blanca y gris del sistema nervioso luego de la aparición de los vasos sanguíneos.
Desarrollo de las placas basales, alares, del techo y del piso
La multiplicación de los neuroblastos de la capa del manto, a cada lado del tubo neural origina unos engrosamientos en la región ventral y dorsal:
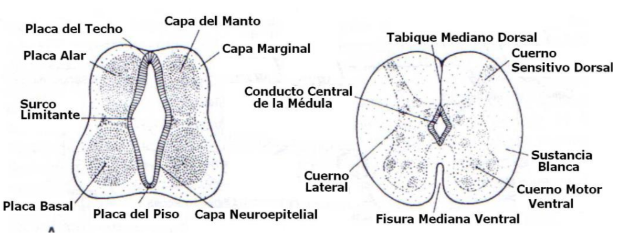
Las regiones dorsal (placa del techo) y ventral (placa del piso) en la línea media del tubo neural no poseen neuroblastos y constituyen vías para fibras nerviosas que cruzan la médula espinal de un lado al otro. Entre los cuernos ventral y dorsal de los segmentos torácicos hasta el segundo o tercero lumbar de la médula espinal se acumulan neuronas que formarán el cuerno lateral o intermedia, que contiene neuronas del sistema nervioso autónomo.
3. Migración Neuronal
Concluido el periodo mitótico, ocurre el fenómeno de migración neuronal, mecanismo que llevará los cuerpos neuronales hasta el sitio donde realizarán sus funciones definitivas. Se ha postulado que esto ocurre con la participación de glías especializadas, formadas por los glioblastos, que son las llamadas células guiadoras (células dianas o guías). La velocidad de migración es lenta (0,001 mm x 24hrs).
En la migración lo primero que ocurre es que el neuroblasto adyacente a la capa ependimaria se adosa a la célula guía y, más tarde, a través de movimientos ameboídeos, éste se desplaza a su lugar definitivo. Éste puede ser en la capa del manto o en la marginal. Por lo tanto, la ubicación de las neuronas en las diferentes capas del cerebro y cerebelo, está dada por las células gliales que están guiando el proceso. Cuando la neurona se contacta con la membrana glial, la célula deja de proliferar y extiende su proceso. La neurona conserva su adhesión mediante una serie de proteínas especialmente la astrotactina.
Las células diana salen de la zona de producción, y tienen largas proyecciones hacia la superficie del órgano, lo que permite que la última neurona en formarse sea la última en llegar a tomar su lugar, esto significa que la formación de neuronas es de profundidad a superficie. Y cuando una célula queda ubicada en mala posición, sufre apoptosis.
4. Agregación Neuronal
Concluida la migración sobreviene el proceso de agregación, fenómeno que determina que algunos neuroblastos permanezcan juntos y formen núcleos o estratos celulares, en tanto que otros se separan y se asocian, estableciendo contactos y relaciones diferentes. La agregación se produce gracias a la participación de MAC como la E- cadherina y la N-cadherina, ubicadas en la superficie de los neuroblastos, que permiten el reconocimiento de las neuronas entre sí. La agregación se ha comprobado a través de experimentos de desagregación celular, que consisten en desagrupar células y observar, obteniendo como resultado que lentamente se acercaban y se volvían a agregar formando un núcleo, por lo tanto existe un “algo” que permite el reconocimiento de ellas entre sí para finalmente formar estos núcleos.
5. Diferenciación Neuronal
Mecanismo por el cual cada neurona adquiere las características morfológicas propias y los contactos sinápticos específicos que las diferencian entre sí. Uno de los factores que regula la diferenciación es el factor de crecimiento nervioso, capaz de producir modificaciones en la morfología celular y en la dirección que siguen estas prolongaciones, es decir, cómo se forman los contactos, cómo se forman las sinapsis, qué neuronas se unen con otras y cuáles no, etc.
Trofismo
Durante el desarrollo del sistema nervioso, se generan más neuronas de las que existen en el adulto. De hecho, más de un 50% de las neuronas en desarrollo, mueren antes de entrar en funcionamiento. Por ejemplo, más de la mitad de las motoneuronas inferiores que sinaptan con músculo esquelético mueren a las pocas horas después de establecida la unión. Esta muerte es resultado de una especie de competencia entre las neuronas por captar las cantidades limitadas de factor neurotrófico liberado por las células musculares, lo que ocasiona una muerte celular programada de las neuronas que no captan lo suficiente. Este es un medio eficaz para ajustar el número de neuronas al número de células efectoras que inervarán.
El agente neurotrófico más caracterizado es el factor de crecimiento neural o NGF (Nerve Growth Factor). Es un elemento esencial para las neuronas sensitivas y simpáticas, ya que es un factor estabilizador de sus sinapsis, es capaz de estimular y conducir el crecimiento y regeneración de sus axones, para así ajustar el suministro de inervación a las necesidades de las células blanco; en efecto, la administración de anticuerpos anti-NGF en un ratón con su sistema nervioso en desarrollo provocó la muerte a todas las neuronas simpáticas y sensitivas.
El NGF es producido por células inervadas por neuronas dependientes de NGF. Luego de ocurrida la muerte de las neuronas sobrantes, el NGF es importante en la mantención de la densidad de inervación ya que controla la cantidad de terminales axonales. El NGF alcanza las neuronas en las terminales axonales y avanza por transporte axonal retrógrado hacia el cuerpo celular para ejercer sus efectos.
El tejido mesenquimático (esclerotoma) que rodea el tubo neural se condensa para formar la meninge primitiva, que originará la duramadre. A esta meninge primitiva se le agregan células provenientes de las crestas neurales para formar la capa interna denominada leptomeninges (aracnoides y piamadre).
Al unirse los espacios llenos de líquidos que existen entre las leptomeninges, se forma el espacio subaracnoídeo. El origen de la aracnoides y piamadre a partir de una capa única explica la existencia de las trabéculas aracnoideas que existen entre ellas.
Como resultado del desarrollo del aparato locomotor durante el 4º mes, además de la adición de neuronas motoras y sensitivas, la médula espinal se ensancha en las regiones cervical y lumbar formando los engrosamientos cervical y lumbar.
Al 3º mes, la médula espinal se extiende a lo largo del canal vertebral del embrión y los nervios espinales atraviesan los agujeros intervertebrales a nivel de su origen. Poco después, la columna vertebral y la duramadre se alargan más rápido que el tubo neural ocasionando que el extremo terminal de la médula se desplace a niveles más altos.
A los 6 meses de vida intrauterina alcanza la 1ª vértebra sacra, y ya en el neonato su extremo está a nivel de L3. Debido a este crecimiento desproporcionado, los nervios raquídeos tienen una dirección oblicua desde su segmento de origen en la médula espinal hasta el nivel correspondiente de la columna a nivel coccígeo.
En el adulto, la médula espinal termina a nivel L2 (esta es una medida promedio, ya que el extremo medular puede estar tan alto como T12 o tan bajo como borde superior de L3). Una prolongación filiforme de la piamadre forma el filum terminale que se adosa al periostio de la 1ª vértebra coccígea y señala la línea de regresión de la médula espinal embrionaria. Las fibras nerviosas bajo el extremo inferior de la médula espinal forman la cauda equina, cuya denominación se debe a su semejanza a la cola de caballo. Cuando se extrae líquido cerebro espinal por una punción lumbar, la aguja se introduce en un nivel lumbar bajo respetando así el extremo terminal de la médula espinal.
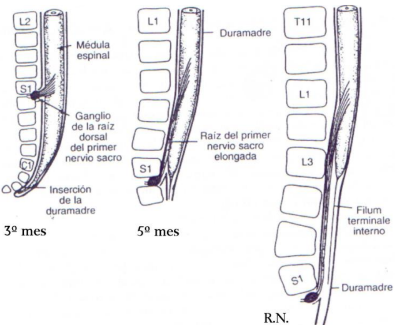
Resumen: falso ascenso de la médula
En los recién nacidos el cono medular está a nivel de L3; es por esto que se habla de un falso ascenso de la médula durante el crecimiento. En un principio, la médula espinal está a la altura de Co1, al quinto mes se encuentra a la altura de S1, en el recién nacido termina a la altura de L3 y en el adulto entre L1 y L2, como ya fue mencionado.
En realidad, no es que la médula ascienda, lo que realmente ocurre es que los segmentos óseos (columna vertebral), se desarrollan más rápido que el tejido nervioso, es decir, tienen un ritmo de crecimiento más rápido que la médula espinal.
Neuroanatomía UFRO by Camila Díaz is licensed under CC BY-NC-SA 4.0![]()
![]()
![]()
![]()