
La médula espinal hacia arriba se continúa con el encéfalo, específicamente la médula oblonga y hacia abajo termina en un ahusamiento propio de la médula espinal, denominado cono medular, cuyo vértice en el adulto se ubica a nivel de la 1ª o 2ª vértebra lumbar.
De la médula espinal se originan 31 pares de nervios espinales, los cuales salen por el respectivo agujero intervertebral. Si bien las raíces de los nervios espinales cervicales corren horizontalmente, las raíces de los otros nervios espinales hacen su recorrido cada vez más oblicuo y vertical mientras más bajo sea segmento medular, esto debido al crecimiento diferencial vertebro-medular (falso ascenso de la médula).
Es así como a las raíces espinales correspondientes a los segmentos lumbares, sacros y coccígeos, que corren verticalmente bajo el cono medular, se les denomina cauda equina por su notable parecido a una «cola de caballo».
La médula esta constituida por segmentos medulares, que corresponden a un área de la médula espinal desde donde se originan las raíces de un nervio espinal, en otras palabras, es toda el área que abarca el origen del nervio, por lo tanto, existen 31 segmentos medulares. Esta organización corresponde a vestigios de la segmentación o metamería del organismo.
Debido a sus relaciones con las siguientes estructuras, la médula puede permanecer fija en su lugar:
La médula espinal externamente es blanquecina, debido a que superficialmente está compuesta de fibras nerviosas mielinizadas. Su longitud es de 45 cms en el hombre y 41-42 cms en la mujer (mientras que la longitud de la columna vertebral es de aproximadamente 73 cms). Tiene forma cilíndrica, ligeramente aplanada en sentido anteroposterior. Su espesor no es uniforme, el promedio es de 0.8 a 1.0 cm. y presenta 2 engrosamientos, llamados Intumescencias:
Bajo el cono medular y hasta la terminación del saco dural en el borde inferior de S2, el espacio subaracnoideo sólo contiene la cauda equina y el filum terminal flotando en LCE Por tal razón, esta es la zona (debajo de L2) con menor riesgo para efectuar una punción lumbar. Al hacerla más arriba, puede provocar una lesión medular, al puncionar la médula.
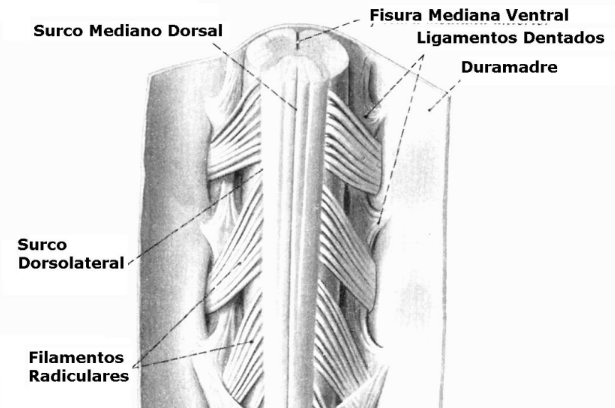
Al desprender las meninges que envuelven la médula espinal, se observan fisuras (profundas) y surcos en la superficie medular:
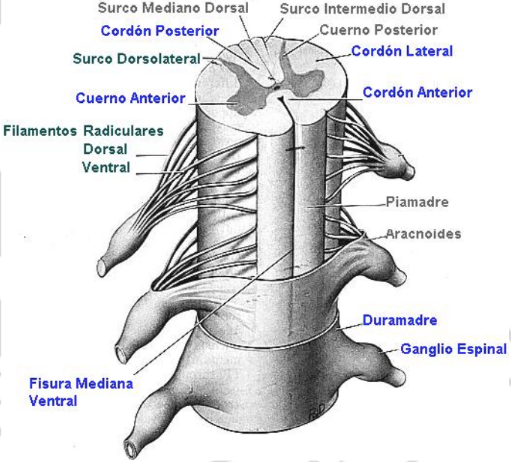
Con propósitos descriptivos, se ha dividido cada mitad de la médula espinal en tres cordones (columnas o funículos) de sustancia blanca, tomando como límites los diferentes surcos y fisuras:
Al observar la sustancia gris, podemos observar que no es igual a lo largo de la médula.
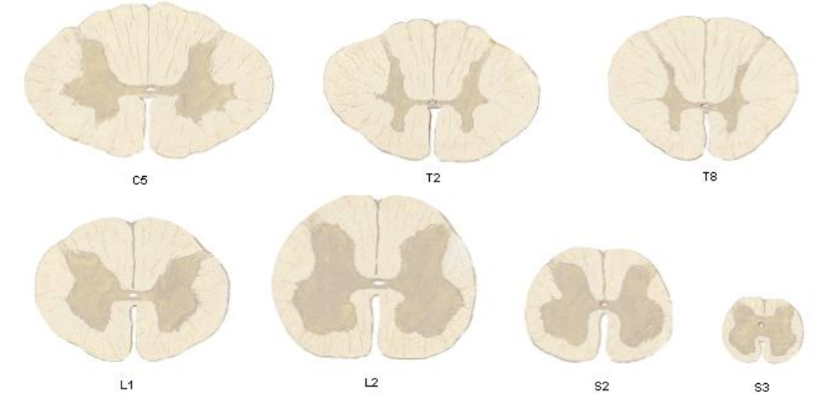
La cantidad de sustancia gris presente en los diferentes niveles medulares depende de la cantidad de músculos inervados por ese nivel. Por tal motivo, en los segmentos torácicos los cuernos ventrales son delgados y pequeños, mientras que en los segmentos cervicales inferiores y lumbosacros son gruesos, ya que contienen las neuronas motoras asociadas a los plexos braquiales y lumbosacros. En los vértices de los cuernos posteriores de estos últimos segmentos es posible observar un área de sustancia gris de apariencia transparente denominada sustancia gelatinosa, que también está presente en segmentos torácicos pero menos aparente debido a su escasez.
Sustancia gris en los diferentes niveles medulares
A nivel cervical, existe un cuerno ventral bastante aumentado, debido a la presencia de gran cantidad de neuronas motoras que controlan los músculos del miembro superior. En la parte lateral de la base del cuerno posterior, por ejemplo de los segmentos cervicales superiores, es difícil distinguir la sustancia gris de la blanca debido a que células y fibras nerviosas se encuentran mezcladas, por la formación reticular, que se continúa superiormente como la formación reticular del tronco encefálico.
A nivel torácico tenemos un cuerno ventral pequeño y un cuerno dorsal bastante alargado. Además a este nivel aparece un cuerno lateral, que se extiende de C8 a L2, que representa la sustancia gris intermedia lateral, lugar donde en el embrión estaba el surco limitante, por lo que corresponde a una zona visceral, viscero-motriz, donde se ubica el cuerpo de la primera neurona simpática, cuyos axones salen por la raíz ventral de los nervios espinales.
A nivel lumbar vuelve a crecer el cuerno ventral, debido a que hay mayor número de neuronas motoras para el control del miembro inferior.
A nivel sacro el cuerno ventral y el dorsal son de gran tamaño lumbar. A nivel de S2, S3 y S4, aparece un cuerno lateral, que contiene el soma de la primera neurona parasimpática, destinada al control de las vísceras de la excavación pélvica. A nivel sacro, los cordones tienen menor cantidad de fibras que a nivel cervical, debido a que la cantidad de fibras, conforme se va ascendiendo, va aumentando.
Topografía vértebro-medular
Es importante conocer la relación entre los segmentos medulares y las vértebras, especialmente para la localización de lesiones.
En un corte transversal se puede observar que la médula espinal consta de una región central, denominada sustancia gris, y una región periférica de aspecto blanquecino denominada sustancia blanca.
La sustancia gris tiene la forma de una “H”, en su parte media está el canal central de la médula (ex-tubo neural); por lo tanto, la sustancia gris que está paralela al canal central, se denomina sustancia gris intermedia, la más cercana es la sustancia gris intermedia central, y el resto es sustancia gris intermedia lateral.
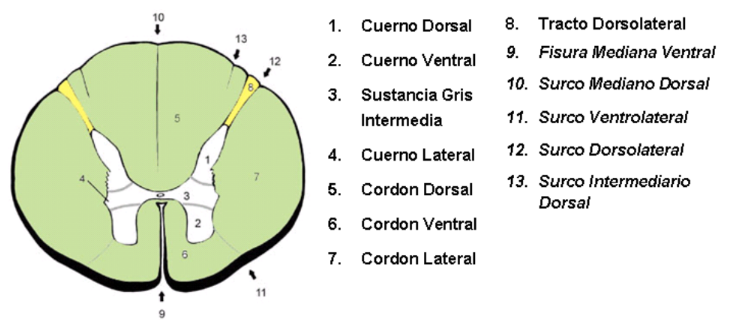
La sustancia gris posee 4 cuernos: 2 ventrales y 2 dorsales.
Cada cuerno ventral, topográficamente, tiene lo que se llama una cabeza y una base; y cada cuerno dorsal tiene una base, un cuello y una cabeza. Esto es muy importante para relacionarlo posteriormente con aspectos funcionales.
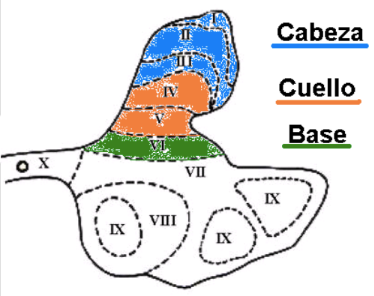
La disposición tridimensional de los cuernos ventrales y dorsales conforma verdaderas columnas que recorren la médula espinal, constituyendo las columnas grises ventrales y dorsales.
Los cuernos dorsales, funcionalmente son sensitivos, están formados por neuronas sensitivas que reciben los impulsos que llegan por las raíces posteriores. Los cuernos ventrales, funcionalmente son somato-motores, están constituidas por neuronas motoras cuyos axones salen por las raíces anteriores. Las columnas grises ventrales y dorsales de cada lado se encuentran unidas por una banda transversal de sustancia gris, denominada comisura gris.
Justo en medio de la comisura gris se encuentra el canal central de la médula espinal, que es un pequeño conducto, lleno de líquido cerebro-espinal (LCE), que recorre completamente a la médula espinal. Este conducto suele ser apenas visible o permanecer ocluido con el epitelio cilíndrico ciliado que lo recubre (epéndimo).
El canal central de la médula espinal se continúa hacia arriba con el canal central de la mitad caudal de la médula oblonga, para luego abrirse paso a la cavidad del cuarto ventrículo. En el extremo inferior de la médula espinal puede formar una dilatación, que se prolonga hacia el filum terminal, denominada ventrículo terminal. El canal central sirve de referencia para dividir la comisura gris en dos mitades en sentido anteroposterior, la comisura gris anterior y la comisura gris posterior. Entre la comisura gris anterior y la fisura mediana anterior existe una banda transversal de sustancia blanca, que comunica ambos cordones anteriores, denominada comisura blanca anterior.
Citoarquitectura de la sustancia gris
Si analizamos la citoarquitectura de la sustancia gris, observaremos neuronas de gran tamaño, y de tamaño pequeño, neuronas de axón largo y de axón corto, teniendo diferentes clasificaciones.
Cuerno ventral:
En el cuerno ventral se ubican principalmente neuronas motoras grandes o alfa-motoneuronas, de forma estrellada y axón largo (neuronas Golgi-1), que pueden ser:
Además existen unas neuronas multipolares pequeñas (10 – 20 um) llamadas gama–motoneuronas y otras de axón corto, ubicadas en la porción medial del cuerno ventral, denominadas neuronas inhibitorias o de Renshaw, que inhiben tanto a las alfa como gama motoneuronas.
Las funciones de las neuronas que se encuentran en el cuerno ventral son:
Sustancia Gris Intermedia:
Está localizada entre los cuernos ventrales y dorsales. Contiene principalmente neuronas de asociación.
Cuerno Dorsal:
Tiene mayoritariamente neuronas receptoras y neuronas de axón corto, que son estimuladas principalmente por impulsos que ingresan a la médula a través de las raíces posteriores, donde se encuentra ubicado el ganglio espinal, que tiene neuronas pseudomonopolares.
Las neuronas pseudomonopolares o células en “T”, tienen 2 prolongaciones, una de ellas va hacia la periferia, y la otra penetra por la raíz posterior a la médula. Su función es transportar el estímulo desde el receptor hacia el cuerno dorsal de la médula.
Por todo lo anterior podemos decir que el cuerno dorsal corresponde a la zona sensitiva de la médula espinal y además, que estaría actuando como un filtro sensorial de estímulos, dejando pasar sólo a algunos a los niveles más altos, mientras que a otros los inhibe o anula.
El cuerno dorsal desde el punto de vista didáctico se puede dividir en:
Cuerno lateral:
Este cuerno los encontramos solamente en 2 zonas de la médula:

Por lo anterior, en la zona donde se presentan las intumescencias, encontramos 6 núcleos en el cuerno ventral de la médula, mientras que en la zona situada entre las dos intumescencias (médula torácica) encontramos sólo dos núcleos:
Entre los grupos mediales y laterales también es posible observar unos “núcleos centrales” que se intercalan.
Núcleos de los cuernos dorsales:
Se disponen en un sentido más bien dorso ventral, alineándose de la siguiente forma:

La médula espinal, al igual que el encéfalo, está envuelta por las meninges: duramadre, aracnoides y piamadre.

Duramadre:
Es la meninge más externa. Es una membrana fuerte, densa y fibrosa que envuelve la médula espinal y la cauda equina. Tiene un color blanco nacarado. Se continúa a través del foramen magno con la duramadre que recubre el encéfalo. En la parte inferior de la médula espinal forma el fondo de saco dural que se extiende hasta S2. Bajo esta vértebra sacra, la duramadre se continúa rodeando al filum terminal y su unión forma el ligamento coccígeo que se inserta en el cóccix. Se continúa en los forámenes intervertebrales, rodeando a los nervios espinales, continuándose a su vez con el epineuro. Está separada de las paredes del canal vertebral por el espacio epidural.
Espacio epidural (o extradural o peridural):
Este espacio es ocupado por tejido conjuntivo laxo, grasa peridural y por el plexo venoso vertebral interno. Este plexo tiene comunicación directa con los senos durales del cráneo, por lo tanto puede ser una vía de diseminación de infecciones, émbolos o células cancerosas desde la pelvis hacia el cerebro. Existe sólo en el canal vertebral, en el cráneo no existe. Protege a la médula de posibles daños producto de los movimientos de la columna.
El espacio epidural es de enorme importancia en clínica, ya que bajo L2 se pueden depositar anestésicos locales (anestesia epidural) que permiten intervenciones quirúrgicas, en general, bajo el ombligo. Es muy usada la anestesia regional sobre todo en el ámbito obstétrico. También se puede extraer líquido cerebroespinal con fines analíticos o para medir presión.
Aracnoides:
Es una membrana delgada e impermeable que recubre totalmente la médula espinal. Se ubica entre la piamadre (que es la membrana más profunda), y la duramadre. Envía prolongaciones en forma de tela de araña hacia la siguiente capa (piamadre), constituyendo el espacio subaracnoideo, por el cual circula el líquido cerebroespinal, que rodea a la médula en toda su extensión, y tiene comunicación con el encéfalo. Se encuentra separada de la duramadre sólo por un líquido.
Espacio subdural:
Este espacio es un “espacio virtual”, ya que, sólo presenta una pequeña cantidad de líquido cerebro espinal, que permite el deslizamiento entre la duramadre y la aracnoides. Es importante en el cerebro, pues cuando hay traumatismos craneanos puede haber ruptura de las arterias meníngeas que circulan por ese lugar, formándose así los llamados hematomas subdurales, que comprometen de forma importante la salud del paciente.
Espacio subaracnoideo:
Este espacio es real, rodea todo el encéfalo y prosigue inferiormente, a través del foramen magno hasta el borde inferior de S2, en donde la duramadre y la aracnoides se fusionan con el filum terminal, no dejando espacio alguno.Su importancia radica en que contiene el líquido cerebroespinal. El espacio subaracnoideo es atravesado por finas trabéculas aracnoideas que se unen a la piamadre.
Piamadre
Es una capa única y delgada, de carácter vascular, que se adosa íntimamente a la médula espinal, es capaz de introducirse en todas las fisuras y surcos. Se puede dividir en dos capas: la más externa o epipia, que está en contacto con el líquido cerebroespinal, siendo la más importante, por circular en ella los vasos sobre la médula espinal; y la más interna que está adosada a la médula. La piamadre envía prolongaciones hacia la duramadre en forma de dientes de sierra llamadas ligamentos dentados.
Los ligamentos dentados se ubican desde el foramen magno a L1. Por las caras laterales de la médula, a igual distancia entre las raíces posteriores y anteriores de los nervios espinales, éstas 21 extensiones membranosas puntiformes de la piamadre, van a insertarse firmemente a la cara interna de la duramadre y aracnoides. Son utilizados como puntos de referencia para procedimientos quirúrgicos. Además facilitan la suspensión de la médula espinal justo en medio del saco dural. En tal función también participan la continuidad con el tronco encefálico, la presión ejercida por el líquido cerebro espinal y el filum terminal.
Neuroanatomía UFRO by Camila Díaz is licensed under CC BY-NC-SA 4.0![]()
![]()
![]()
![]()